Thread Truncated (Cap Enforced)
Only the first 20 tweets are unrolled into slides to ensure reliable PDF exporting and high server performance.
Canvas & Ratio
Choose your destination platform format
Layout Template
Choose a content structure for your slides
Preset Themes
Typography & Sizing
Brand Kit Customization
AGENCYConfigure brand assets for headers & footers
Outro Slide CTA
Customize your closing call-to-action slide
Background Pattern
Build Your Carousel
Drag and drop any post card below onto a slide, or use the quick buttons to insert content/images instantly!

Could the folding of synthetic gene circuits in 3D shape how genes are expressed? Today @ScienceMagazine we report on the role of gene syntax in shaping feedback between transcriptional activity and genome folding for advanced circuit design🧵 (1/n)

What’s gene syntax? Gene syntax is the relative order and orientation of genes. Just as word order in a sentence can alter emphasis and even meaning, we predicted that the ordering of genes can change how genes are expressed, amplifying or dampening expression. (2/n)
You can read the full thread on our predictions from the biophysical modeling here. Or continue on for the TL/DR and new experimental results! (3/n) <a target="_blank" href="https://x.com/GallowayLabMIT/status/1582449416534773760" color="blue">x.com/GallowayLabMIT…</a>
TL/DR: As RNAP transcribes, it generates a wave of overwound DNA (+ supercoiling) ahead & leaves a wake of underwound DNA (- supercoiling). Accumulated supercoiling both reshapes the RNAP binding energy landscape and can stall polymerases, creating supercoiling-mediated feedback! Feedback is an essential mechanism for building stable control systems. But control needs to be both fast and precise for optimal stability. (4/n)

What’s special about supercoiling-mediated feedback? It is FAST, really fast! Because this feedback operates at the speed of supercoiling generation, we have very rapid feedback that impacts subsequent RNAP binding and transcriptional events, coupling adjacent genes. This much faster than most feedback loops which go through multiple processes before feeding back to regulation. (5/n)

But if syntax matters, then shouldn’t we see patterns of syntax enrichment in genomes? Indeed, clues to this hidden context are sprinkled across the genomes of organisms from yeast to humans. The syntax of pairs of genes are not uniformly spaced. Mice and humans show enrichment of divergent pairs at short spacing, whereas yeast enrich for convergent syntax. (6/n)
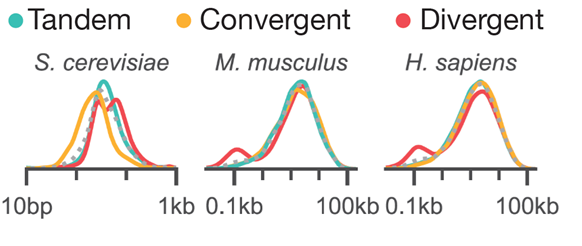
This enrichment is even more striking when examining only the pairs of genes that are BOTH actively transcribed in human iPSCs. Almost all of the co-active pairs are divergent! (7/n)
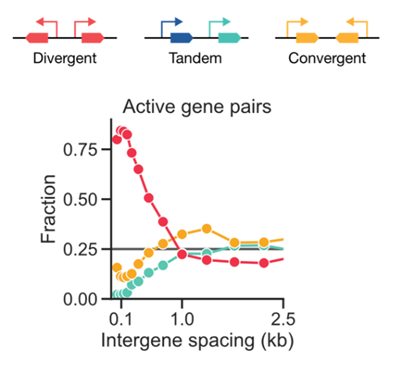
Ok, so it may be that the human genome is using syntax to optimize expression of pairs of genes. Or maybe not. Feynman famously said, “What I cannot create, I do not understand.” What we needed to do next was to build some of these two-gene systems and test the predictions! (8/n)

We had been sharing the modeling work within the synthetic biology community as we explored the predictions, Ross Jones @jonesr18 from Peter Zandstra’s lab @PZandstra reached out to say he was observing some of the position effects we had predicted. (9/n)
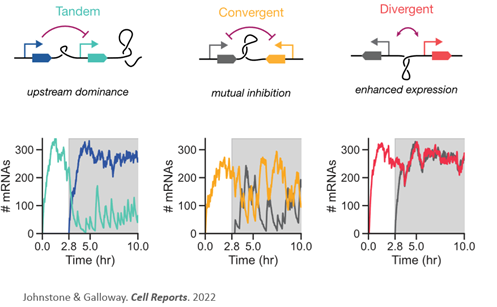
He had put two transcriptional units in tandem and delivered them via PiggyBac to hiPSCs. The upstream gene was very strongly expressed and the downstream gene was weakly expressed, putatively this was the upstream dominance that we had predicted. What struck us was that he had switched the transcriptional units (promoter-FP-PAS) to verify position (not sequence) was determining the bias. Remarkably, the switch led to a 24x fold change in expression from one of the reporters! Woah! That was astonishing. (10/n)

Ok but these are “static promoters”. Maybe there’s some odd coupling that could explain this. What I really want to see is the dynamic response! Turn it on, turn it off! (like Cheat here throwing his lightswitch rave, IYKYK) (11/n)
To answer what happens when we turn on the adjacent gene, we build two-gene systems with a constitutive promoter (should be constant) and an inducible promoter (we can turn it on/off with a small molecule, dox). We randomly integrated these two gene systems with different syntaxes into human cells (293Ts). So what happens when we add dox? (12/n)

Adding dox reduces expression from the downstream gene in tandem. For the convergent, the expression is very stable (as are the proteins we use to measure!). For the divergent, we see a very strong, dox-dependent increase in the reporter. This is the predicted amplification! 🪩(13/n)

And these differences are very clearly visible, they are not mild effects! While the previous data was normalized, you can also see how poorly the convergent expresses very, very low but above negative controls) (14/n)

Ok, but is this syntax-effect uniform, suggesting a broad shift in transcription rates? or could this be some subpopulation effect? Looking at both protein and RNA (using Flow-FISH) we saw unimodal shifts in expression indicating changes in protein levels likely reflect broad, syntax-specific changes in transcription rate upon induction! (15/n)
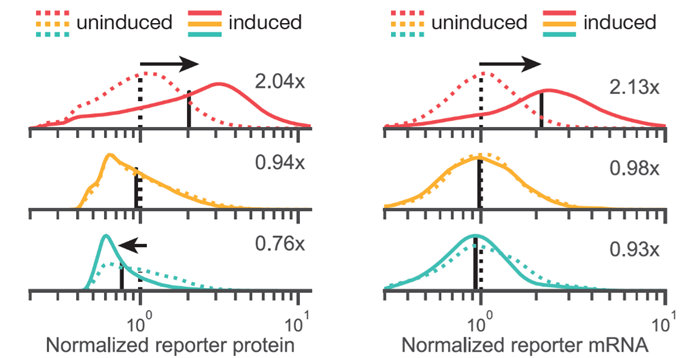
Ok, but maybe this is some epigenetic shift? If it’s supercoiling, you should be able to switch it on and off and see the system relax to the initial state. Yes, indeed! (16/n)
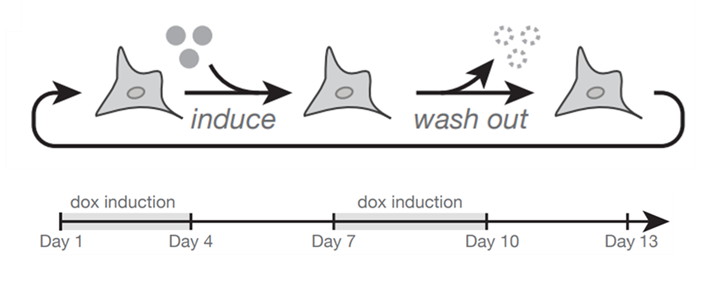
To test that idea, we ran the experiment over 13 days, pulsing expression on and off every 3 days. And indeed, the inducible gene goes up and down with induction for all syntaxes (top graph), but the reporter shows syntax-specific patterns. Expression of the reporter amplifies in divergent syntax, stays constant with convergent, and is reduced when the reporter is downstream tandem. (17/n)

So the coupling pattens are reversible and dependent on the activity of the adjacent gene, just as predicted by our model of supercoiling-mediated feedback! But…are we actually changing supercoiling? Does induction of expression change chromatin folding? Or is this just a nice campfire story o_O (18/n)

To directly examine chromatin folding and supercoiling, we wanted to place our circuit precisely within the genome to ensure that comparisons were as controlled and quantitative as possible. To measure folding at the high resolution needed for gene circuits, we also needed to integrate the circuits at both alleles so the lines were homozygous for each circuit (e.g. each syntax site-specifically integrated at both alleles). (18/n)

if you do any type of mammalian cell engineering, you would know that is a tall order! Fortunately, @A_BlanchAsensio from the Mummery and Davis labs has been working with us on a solution. STRAIGHT-IN DUAL! Using STRAIGHT-IN Dual we could integrate our circuits homozygously AND remove flanking regions which support rapid integration but could confound our measurements. (20/n)
