@GallowayLabMIT: Could the folding of synthetic...
@GallowayLabMIT
7 views
May 02, 2026
Advertisement
1
Could the folding of synthetic gene circuits in 3D shape how genes are expressed? Today @ScienceMagazine we report on the role of gene syntax in shaping feedback between transcriptional activity and genome folding for advanced circuit design🧵 (1/n)

2
What’s gene syntax? Gene syntax is the relative order and orientation of genes. Just as word order in a sentence can alter emphasis and even meaning, we predicted that the ordering of genes can change how genes are expressed, amplifying or dampening expression. (2/n)
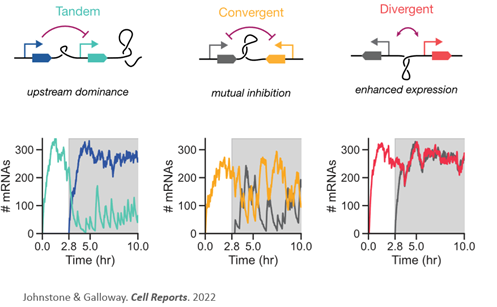
3
You can read the full thread on our predictions from the biophysical modeling here. Or continue on for the TL/DR and new experimental results! (3/n)
View Tweet
4
TL/DR: As RNAP transcribes, it generates a wave of overwound DNA (+ supercoiling) ahead & leaves a wake of underwound DNA (- supercoiling). Accumulated supercoiling both reshapes the RNAP binding energy landscape and can stall polymerases, creating supercoiling-mediated feedback! Feedback is an essential mechanism for building stable control systems. But control needs to be both fast and precise for optimal stability. (4/n)

5
What’s special about supercoiling-mediated feedback? It is FAST, really fast! Because this feedback operates at the speed of supercoiling generation, we have very rapid feedback that impacts subsequent RNAP binding and transcriptional events, coupling adjacent genes. This much faster than most feedback loops which go through multiple processes before feeding back to regulation. (5/n)
6
But if syntax matters, then shouldn’t we see patterns of syntax enrichment in genomes? Indeed, clues to this hidden context are sprinkled across the genomes of organisms from yeast to humans. The syntax of pairs of genes are not uniformly spaced. Mice and humans show enrichment of divergent pairs at short spacing, whereas yeast enrich for convergent syntax. (6/n)
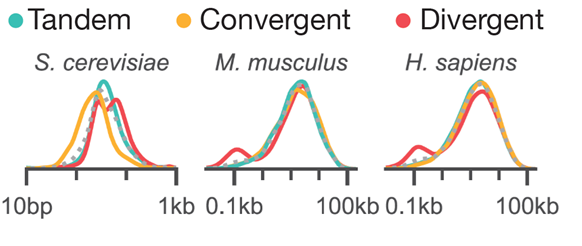
7
This enrichment is even more striking when examining only the pairs of genes that are BOTH actively transcribed in human iPSCs. Almost all of the co-active pairs are divergent! (7/n)
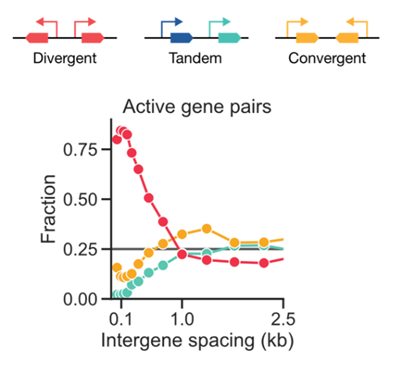
8
Ok, so it may be that the human genome is using syntax to optimize expression of pairs of genes. Or maybe not. Feynman famously said, “What I cannot create, I do not understand.” What we needed to do next was to build some of these two-gene systems and test the predictions! (8/n)

9
We had been sharing the modeling work within the synthetic biology community as we explored the predictions, Ross Jones @jonesr18 from Peter Zandstra’s lab @PZandstra reached out to say he was observing some of the position effects we had predicted. (9/n)
10
He had put two transcriptional units in tandem and delivered them via PiggyBac to hiPSCs. The upstream gene was very strongly expressed and the downstream gene was weakly expressed, putatively this was the upstream dominance that we had predicted. What struck us was that he had switched the transcriptional units (promoter-FP-PAS) to verify position (not sequence) was determining the bias. Remarkably, the switch led to a 24x fold change in expression from one of the reporters! Woah! That was astonishing. (10/n)
11
Ok but these are “static promoters”. Maybe there’s some odd coupling that could explain this. What I really want to see is the dynamic response! Turn it on, turn it off! (like Cheat here throwing his lightswitch rave, IYKYK) (11/n)
12
To answer what happens when we turn on the adjacent gene, we build two-gene systems with a constitutive promoter (should be constant) and an inducible promoter (we can turn it on/off with a small molecule, dox). We randomly integrated these two gene systems with different syntaxes into human cells (293Ts). So what happens when we add dox? (12/n)

13
Adding dox reduces expression from the downstream gene in tandem. For the convergent, the expression is very stable (as are the proteins we use to measure!). For the divergent, we see a very strong, dox-dependent increase in the reporter. This is the predicted amplification! 🪩(13/n)
14
And these differences are very clearly visible, they are not mild effects! While the previous data was normalized, you can also see how poorly the convergent expresses very, very low but above negative controls) (14/n)

15
Ok, but is this syntax-effect uniform, suggesting a broad shift in transcription rates? or could this be some subpopulation effect? Looking at both protein and RNA (using Flow-FISH) we saw unimodal shifts in expression indicating changes in protein levels likely reflect broad, syntax-specific changes in transcription rate upon induction! (15/n)
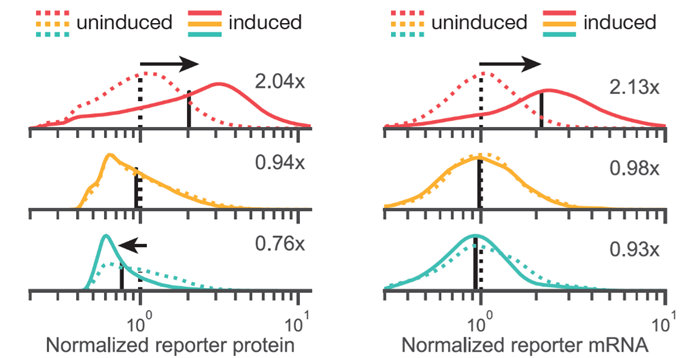
16
Ok, but maybe this is some epigenetic shift? If it’s supercoiling, you should be able to switch it on and off and see the system relax to the initial state. Yes, indeed! (16/n)
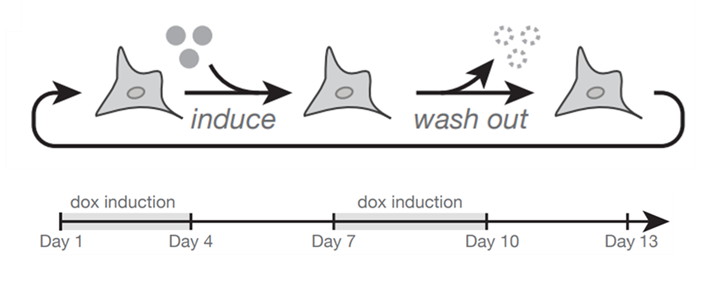
17
To test that idea, we ran the experiment over 13 days, pulsing expression on and off every 3 days. And indeed, the inducible gene goes up and down with induction for all syntaxes (top graph), but the reporter shows syntax-specific patterns. Expression of the reporter amplifies in divergent syntax, stays constant with convergent, and is reduced when the reporter is downstream tandem. (17/n)

18
So the coupling pattens are reversible and dependent on the activity of the adjacent gene, just as predicted by our model of supercoiling-mediated feedback! But…are we actually changing supercoiling? Does induction of expression change chromatin folding? Or is this just a nice campfire story o_O (18/n)

19
To directly examine chromatin folding and supercoiling, we wanted to place our circuit precisely within the genome to ensure that comparisons were as controlled and quantitative as possible. To measure folding at the high resolution needed for gene circuits, we also needed to integrate the circuits at both alleles so the lines were homozygous for each circuit (e.g. each syntax site-specifically integrated at both alleles). (18/n)

20
if you do any type of mammalian cell engineering, you would know that is a tall order! Fortunately, @A_BlanchAsensio from the Mummery and Davis labs has been working with us on a solution. STRAIGHT-IN DUAL! Using STRAIGHT-IN Dual we could integrate our circuits homozygously AND remove flanking regions which support rapid integration but could confound our measurements. (20/n)
21
So once you can integrate these, we verified the patterns of expression were preserved in hiPSCs with site-specific integration and again we saw upstream dominance, and amplified divergent expression!
We couldn’t have done this without the STRAIGHT-IN Dual lines and platform. for more on STAIGHT-IN DUAL which was just published today at Nature Biomedical Engineering, see tweetorial here! (21/n)
We couldn’t have done this without the STRAIGHT-IN Dual lines and platform. for more on STAIGHT-IN DUAL which was just published today at Nature Biomedical Engineering, see tweetorial here! (21/n)
View Tweet
22
Ok so now that we have our circuits precisely placed in the genome. How can we see if induction reshapes folding and patterns are mediated by DNA supercoiling?
We can measure chromatin folding by assaying contact probability and supercoiling by using a protein called GapR that binds to positively supercoiled DNA. Recent advances in both of these methods were important for us to make these measurements… (22/n)
We can measure chromatin folding by assaying contact probability and supercoiling by using a protein called GapR that binds to positively supercoiled DNA. Recent advances in both of these methods were important for us to make these measurements… (22/n)

23
For contact probability, we used Region Capture Micro-C (RCMC) a methods developed by @Anders_S_Hansen's lab which supports targeted pulldown of a region of interest (figure from Goel, V et al. Nature Genetics. 2023) . Since we are just looking at one locus (CLYBL), we want to spend our sequencing dollars getting high resolution at this locus! And boy did we! ( 23/n)

24
Regions of increased proximity can be seen as these triangles which indicate the things are closer in 3D space than you would expect based on their 1D position. (24/n)

25
We can also look for other structures in these heat maps such as plectonemes, the buckled structures that form as twisted (supercoiled DNA) is converted to writhe (buckled). While loops form corner dots, the plectonemes would be expected to generate larger anti-diagonal scores. (25/n)

26
Looking across the locus, we see the antidiagonal score is very small….until we turn on the adjacent gene! And then wow, a spike in anti-diagonal score right at the locus of integration across all syntaxes! Transcription is reshaping folding! (26/n)
27
Zooming in and comparing the pre- and post-induction, we see syntax specific folding around the promoters. The red regions are the increased contacts and for tandem and divergent the antidiagonal spike is very large! Could this be supercoiling? Could these really be plectonemes? (27/n)

28
To examine that question, we needed to directly look at DNA supercoiling. We used technique called GAP-RUN uses MNase to DNA around regions of positive supercoiling, allowing us to read out these region across the genome. This technique was modified from the original GapR-seq developed by Guo and Laub to measure positive supercoiling in bacteria and yeast. (28/n)
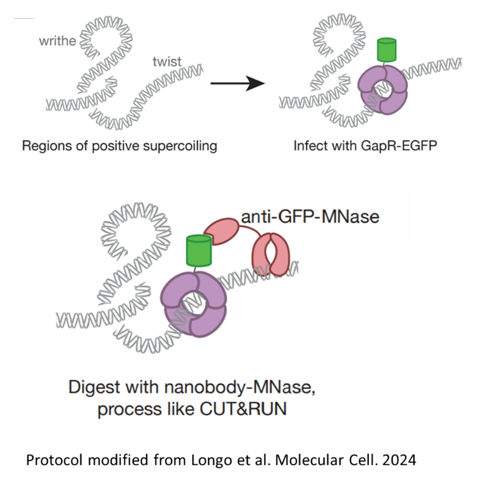
29
Ok, so what do we see? Prior to induction in tandem, we see that positive supercoiling accumulates at the end of the constitutive reporter as expected. But then… (29/n)

30
We turn on the upstream gene and positive supercoiling moves to occlude the constitutive promoter, just as we predicted and in alignment with our observation of upstream dominance. (30/n)

31
See prediction of tandem here. (31/n)
View Tweet
32
But in the divergent, induction results no increase in supercoiling across the promoters. Instead, we see more H3K27ac (red track) at the TSSs as expected. Positive supercoiling accumulates at the end of the genes, just as predicted! (32/n)

33
These patterns largely match our predictions from 2022 (induction shifts positive supercoiling upstream in tandem; supercoiling accumulates at the ends of genes in divergent) but there are differences that need future exploration! (33/n)

34
Looking across other marks at this locus, we can see induction changes the transcription activity, elongating RNA Pol II, and histone marks. Curiously, we do see lots of RNA at the downstream gene in tandem but these transcript appear poorly productive in making the protein, possibly representing readthrough transcripts. (34/n)

35
There’s lots more here to dive into but we are getting our first look at what happens when you turn on gene circuit! Not only are we seeing the first high-resolution image of circuits folding in response to transcription but we can also see how DNA-binding proteins respond to transcription, further supporting a modeling of feedback between transcription, chromatin folding, and gene expression! And we can start to appreciate why syntax matters! 👯 (35/n)
36
Ok so we do see changes in chromatin folding and DNA supercoiling which match our predictions and the syntax-specific patterns of expression, but what can you do with syntax? Can it be used as a design parameter? Can it be useful?... (36/n)
37
Yes, syntax strongly affects expression across delivery modes including lentivirus. Here, in titer-matched viruses, you can see that syntax defines how much induction you can achieve. Notably, the tandem design with the upstream tet and the divergent syntax is the most common in published lentis (by a wide margin!). (37/n)
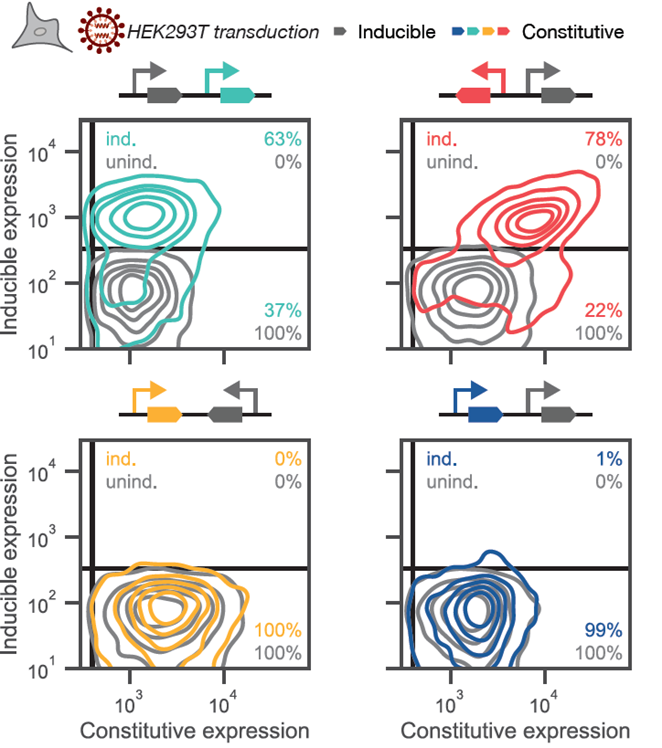
38
Also, the coupling between genes differs by syntax. Divergent syntax couples expression, resulting in a narrower ratio between genes, whereas the tandem allows for a great range of ratios. So syntax can be used to different effects. (38/n)

39
This coupling is similar but slightly different in “all-in-one” vectors where the activator is expressed from the same virus and thus the same locus instead of separately. (39/n)
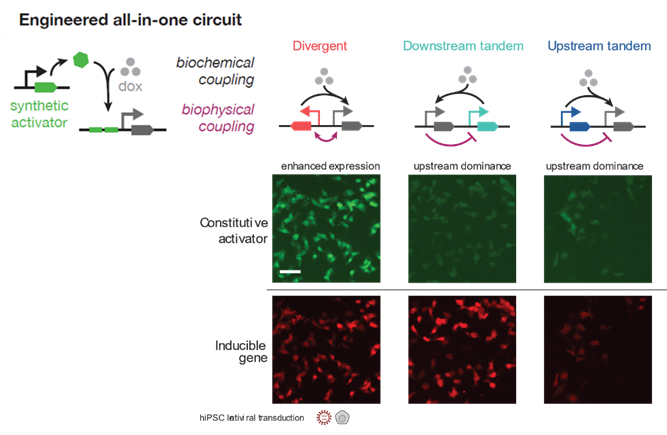
40
Do these couplings change when we increase the distance between the promoters? Not within ~2kb (40/n)

41
What about if we add in insulators? No, insulators exert no significant effect on coupling. Syntax dominates the profiles. (41/n)

42
Do syntax trends persist as the length of the gene increases? Yes, syntax trends are preserved BUT increasing the length reduces expression of the inducible gene…which is interesting. (42/n)

43
Also, in these all and one systems, we noted a curiosity that did not exist for the non-all-in-one (e.g. the separate integration of rtTa). Induction generally increases expression of the constitutive, suggesting some local effect such as TF recruitment, co-factors, condensates, etc. Beyond the mechanism, can syntax be useful beyond reporters? Yes, indeed… (43/n)
44
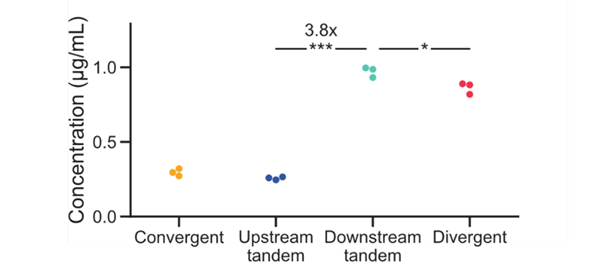
One nice thing about syntax is that you can use it to optimize expression *even if you don’t change parts*! Most of the time people will swap out parts such as promoters to change expression but what if you have limited parts or don’t want to change them? Can this work for optimizing production of biologics? To answer that, we tried producing a difficult-to-produce Yellow Fever antibody discovered by the Dekosky lab. (44/n)

45
Excess light chain is expected to support higher titers of intact mAb. Indeed by ELISA, we saw a 4-fold increase in antibody titers across different syntaxes. (note: all integrated at single copy at the same site in 293Ts). So, yes, even when parts and copy number are constrained, we can use syntax to optimize expression of a biologic! But what about dynamic systems… (45/n)
46
Synthetic biology offers enormous potential in for building sense-and-respond systems and other dynamic circuits such as toggle switches and oscillators for cell and gene therapies. However, accomplishing this vision has been limited by challenges in predictive design including poor choices of syntax (e.g. syntax couples expression in previously unappreciated ways). But with syntax, we also have predictions about how to *improve the design of dynamic systems*. (46/n)

47
Because of the effects of supercoiling-mediated feedback, syntax can generate toggle switches that are biases or balanced in bistability or completely monostable. Syntax can determine if oscillations are coupled or damped. These dynamic behaviors emerge from changes in binding probability. So can we see any evidence of these predicted changes? Even if we see supercoiling as predicted, is there any reason to believe you can design a better dynamic system? So glad you asked!... (47/n)

48
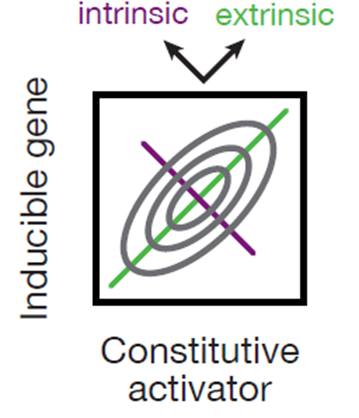
We can most directly see supercoiling-mediadated changes in RNAPII binding probability by looking at intrinsic noise.
What’s intrinsic noise? Intrinsic noise is the variance of expression between genes. If you examine two genes such as the inducible gene and constitutive activator, you will get a distribution of both in a joint plot (shown here). The diagonal is the extrinsic variance which is how the genes scale together and this is often a function of cell size, metabolic activity etc. For example, bigger cells make more of both genes, smaller cells make less of both. (48/n)
What’s intrinsic noise? Intrinsic noise is the variance of expression between genes. If you examine two genes such as the inducible gene and constitutive activator, you will get a distribution of both in a joint plot (shown here). The diagonal is the extrinsic variance which is how the genes scale together and this is often a function of cell size, metabolic activity etc. For example, bigger cells make more of both genes, smaller cells make less of both. (48/n)
49
The antidiagonal dimension measures the correlation between genes, inherently controlling for the other variables (e.g. extrinsic variance). By examining the width of this distribution you get an idea of the intrinsic noise and so what do we see? The divergent distributions are much narrower along the y-axis whereas the tandem is broader across the antidiagonal, just as predicted! (49/n)

50
In quantifying noise, we see that these vectors don’t affect extrinsic variance (which is good!) But we see 3-fold reduction in intrinsic noise in divergent syntax compared to the tandem. This is excellent evidence that syntax biases the probability of co-expression as predicted by our model of supercoiling-mediated feedback! (50/n)

51
To compare across delivery modes and cell types, we can compute the noise ratio. Across 293Ts and hiPSCs, we see that divergent syntax minimizes the noise ratio whether in lenti delivery. Remarkably the noise ratios for divergent syntax of the all-in-one system delivered in lenti or integrated at single copy in landing pads are very similar, suggesting that we don’t need precise, site-specific integration to build systems with minimal intrinsic noise. Indeed, the site-specific tandems perform worse-- by noise ratio -- than the divergent lenti vectors, indicating that syntax is MORE important for reducing noise than controlling for locus of integration! I don’t think we would have expected that apriori. (51/n)

52
As someone who always dreamed of building dynamic genetic control systems, this feels like an inflection point in our understanding and capabilities which have been limited in primary cells!
Thanks @ProfTomEllis 😊for helping articulate how hard this is and @Roth_Lab for noting the timescale! (52/n)
Thanks @ProfTomEllis 😊for helping articulate how hard this is and @Roth_Lab for noting the timescale! (52/n)
53
@ProfTomEllis @Roth_Lab For our own work this has been a huge step forward in forward design, enabling more rapid DBTL cycles and improved single-shot cell engineering that is so much easier and successful than when we started in 2019…so now we can update the meme (54/n)

54
@ProfTomEllis @Roth_Lab Where can get to in the next decade? Something like this. If you were at SynBio 5.0, you may recognize the cat with tophat and monocle. (55/n)

55
Lessons learned: Optimal control comes from working with those that are around you in a symphony, harmonizing requires thoughtful coordination and the right pairing. Those around us really influence how productive and awesome our work can be which is a great transition to recognizing all those that made this work come alive! (somebody queue up Teddy Swims!)(56/n)
56
This project was expertly led by the talented Christopher Johnstone who started on this project in early 2020 just as the pandemic was hitting. Over the first two years, Chris worked to develop the biophysical model that led to our predictions of how syntax could tune supercoiling-mediated feedback and built the experimental systems to test inducible control and then really cranked on getting the genomics assays to test the prediction that transcription induces folding and changes in the patterns of supercoiling. These were all new assays for our lab and he built them from end-to-end! A truly tour-de-force in relentlessly pursuing his hypothesis from cell engineering to genomics! (57/n)

57
Chris was joined by Kasey who first showed that syntax could be used to optimize gene circuits, demonstrating that the divergent substantially outperformed the tandem design. Her leading efforts in using syntax in forward design helped drive the success of many of the papers from our lab in the last year. (58/n)

58
@ProfTomEllis @Roth_Lab We had excellent contributions across the lab and from the Zandstra, Dekosky, Davis, and Mummery labs which helped us really push the limits of the engineering and exploration of genomic circuits. (59/n)

59
@ProfTomEllis @Roth_Lab @kaseyslove @jonesr18 @A_BlanchAsensio @DeonPloessl @emmapeterman_ @science_sneha @PZandstra @MummeryLab Thanks also to our funders including NIH, NSF, AFOSR, Pershing Square and others! In a challenging year, this funding has made a huge difference in our ability to finish the work! (60/n)
60
Thanks to our editor Di Jiang @dijiang319 and the team at Science who helped us with getting helpful, critical feedback on the manuscript. We had great, insightful questions from our reviewers and appreciate their suggestions that did help us nail down the supercoiling mechanism. (61/n)
61
@ProfTomEllis @Roth_Lab @kaseyslove @jonesr18 @A_BlanchAsensio @DeonPloessl @emmapeterman_ @science_sneha @PZandstra @MummeryLab @dijiang319 The vision for this work started eight years ago with just a spark of an idea at mSBW 2018. To see how far we have been able to push this and see supercoiling-mediated feedback come to life has been a wonderous journey that is opening up new roads and adventures… (fin!)